Life at the Cell and Below-Cell Level. The Hidden History of a Fundamental Revolution in Biology
"Dr. Ling is one of the most inventive biochemist I have ever met." |
Chapter 9. Troshin's Sorption Theory for Solute Distribution (p. 43-46) |
|
In 1956, A. S. Troshin
published the Russian version of his monograph,90
which was to be translated into three other languages (German,91
Chinese, and English). The English version bears the title: "Problems of
Cell Permeability."92 Interestingly enough, the contents of
these monographs do not focus on what the title of the book tells us,
permeability, which is a rate process. Instead, they deal primarily with
solute distribution, which in the view of Troshin and
others including myself, represents an equilibrium
phenomenon. A possible interpretation of Troshin's
choice of the title may be as follows: Troshin knew
how the proponents of the membrane (pump) theory—who are by far the majority in
this field—misinterpretated these equilibrium
distribution problems as problems of membrane permeability, but he chose to
refer to these underlying phenomenon as "permeability" problems to be
in touch with a larger audience. In the Preface to the original Russian edition and
reprinted in the English edition of Troshin's book, Nasonov wrote: "We have come to the conclusion that
this theory (the membrane theory) gives a completely false idea of the
structure of the cell and the nature of the substance contained in the protoplasm.
Furthermore, as a result of the apparent simplicity of the scheme it offers in
explanation of the many problematic phenomena, the membrane theory has acquired
great popularity among physiologists and has, in our opinion, induced them to
follow a false line in their theoretical researches."92 p xiii And this is how Troshin
himself introduced his own book: "According to the theory developed by Lepeschkin, Nasonov and Fischer,
and also certain other workers, the greater or lesser permeability of cell for
any substance is to be explained not by the greater or lesser penetration of
the substance through the cell membrane, but by the difference in the
solubility of the substance in the protoplasm and the surrounding aqueous medium
and by the adsorption or chemical binding by the cell colloids of the matter
penetrating the wall."92 p 3 This self-effacing declaration notwithstanding, I
believe that Troshin deserves the lion's share of
credit for the "Sorption Theory" for solute distribution in living cells.
It is true that Moore, Roaf, Fischer, Lepeschkin, Nasonov and others
had pronounced these basic ideas earlier—which is certainly of prime importance
and each of these pioneers should be acknowledged for their priority—but it
was Troshin who transformed these ideas summarily
expressed into quantitative data expressed in a rigorous equation form. To
introduce Troshin's work, I shall begin with the
earlier work of I. Ye. Kamnev. In 1938 Kamnev published in
the Russian-language journal, Archives of Anatomy, Histology and Embryology,
an article entitled, "The permeability for sugars of striated frog
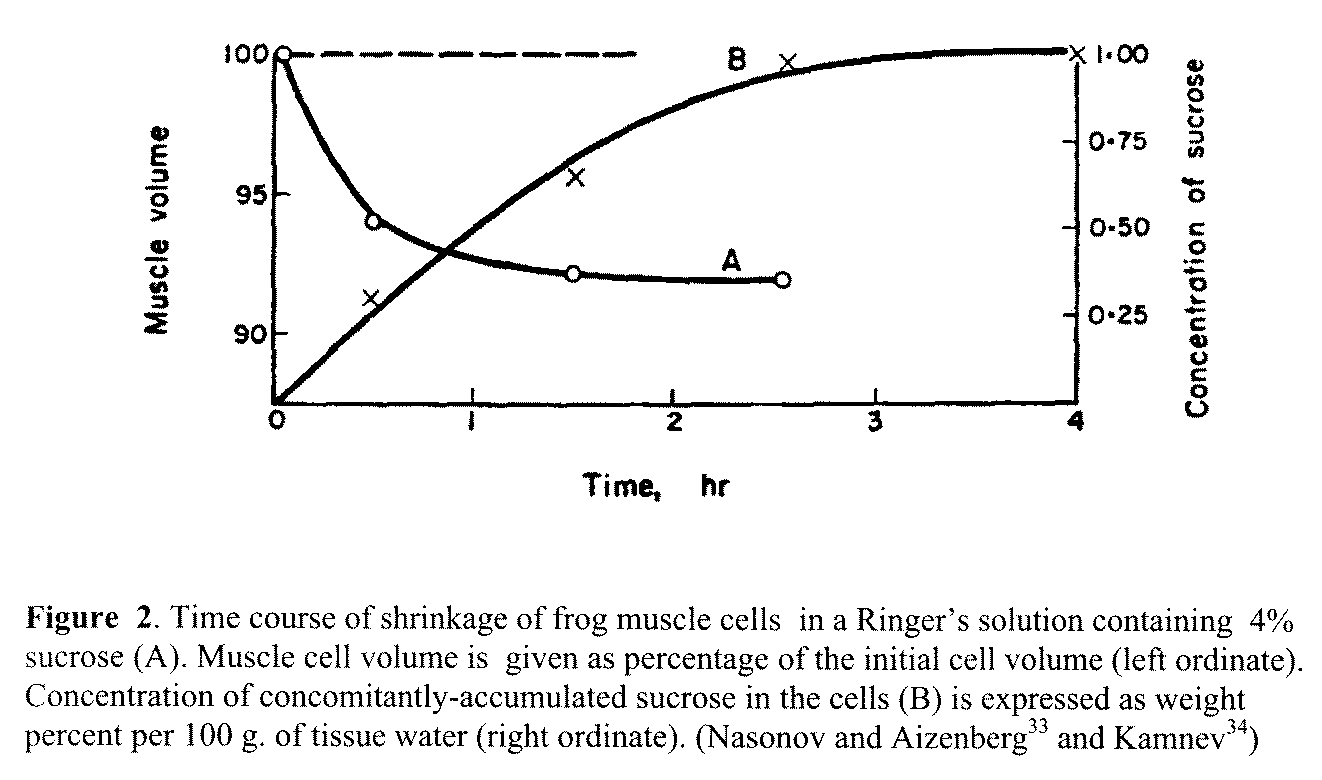
muscle."34 This is a simple but defining paper. When frog
muscles were immersed in a Ringer's solution containing sucrose or galactose, both sugars readily entered the muscle cells and
eventually reached constant levels, which were below their respective levels
in the bathing solutions (Figure 2). In dead cells, the level of sucrose and galactose rose to levels similar to those in the bathing
medium. Kamnev's conclusion that sucrose and galactose
enter the muscle cells rests squarely upon the accuracy of his estimate that
frog sartorius muscle has an extracellular space equal to 9% of its
total muscle weight86 p 114 (and that sucrose or galactose found beyond what is contained in this extracellular
space has entered the muscle cells). Since extracellular space as high as 35%
has been reported in the literature,see
336 p 677 confirmation of the 9% figure is vital to
establishing the validity of Kamnev's conclusion. Work from my own laboratory supports Kamnev. Thus between 1967 and 1975, my associates and I
obtained from five independent methods, four of which were entirely new
(low concentration inulin probe method, 10.3%;
poly-glutamate probe method, 8.9%; single fiber sucrose space method, 9%; Br86
efflux-analysis method, 8.2%; the centrifugation method, 9.4%)49 p 136
an average value of 9.2% ± 0.69% (mean ± S.D.) for the extracellular space of
frog sartorius muscle, which lies close to Kamnev's
9% figure. My evidence that sucrose enters frog muscle cells as shown in Table
1 above support Kamnev's conclusion from a different
angle. Kamnev concluded that the steady level of sugar reached in
the muscle cells does not depend on a membrane mechanism but on the solubility
of these sugars in the muscle sarcoplasm, extending
the idea Martin Fischer first suggested [7]. Kamnev
expressed the belief that the sarcoplasm behaves as
a phase with different solvent properties from water in the surrounding
medium. Ten years after Kamnev's
publication, Troshin resumed this line of study,
indicating that various other nonelectrolytes behaved
just like galactose and sucrose in reaching steady
levels in cell water lower than in the surrounding medium.93 It is
the rapid and seemingly equal rates of entry of these solutes into frog muscle
and other living cells {see [16.6(3.2)]}, which apparently led Troshin's mentor, Nasonov to the
conclusion that there is no cell membrane.86 p 164
(As pointed out earlier, I do not agree with Nasonov
on this specific point. I also believe that to no small measure is this
disagreement a consequence of my access to the powerful radioactive-tracer
technology, apparently unavailable at their times to Nasonov
and Troshin.) Following Bungenberg de
Jong,321 Lepeschkin,324 Duclaux, Guilliermond, Oparin and others,824; 92 p 58 Troshin suggested that living cells are complex
coacervates. Quoting work from Bungenberg de Jong's laboratory and his own, Troshin
showed that, like water in living cells, the water in a simple gelatin
coacervate accommodates various solutes at levels lower than that in the
surrounding medium. Neither Kamnev nor Troshin offered a molecular explanation how water in living
cells differs from normal liquid water, nor a
molecular mechanism why sucrose, galactose have low
solvency in cell water. Nor did they offer an explanation why urea and ethylene
glycol distribute equally across the muscle cell surface [5.1]. On the role of
metabolism on solute distribution, Troshin wrote: "Thanks to metabolism, the sorptional
activity of the protoplasm is maintained in a definite level.....Upon
breakdown of metabolism, this level changes: the solubility of substances in
the protoplasm increases and the binding by cell colloids of some substances is
depressed...."92 pp 373-374 No mechanism was offered why
breakdown of metabolism leads to increased solubility and decreased binding.
Four years earlier, Nasonov concluded his section on
"phase theory of bioelectric potentials and cellular metabolism" in
these words: "energy is evidently necessary for the maintenance of certain
labile chemical compounds (including the structure of proteins)."86 p
202 But he too offered no
explanation of how energy maintains the structure of proteins and other labile
compounds. |
|
|
Разделы книги
Contents (PDF
218 Kb) |
|
|
На страницу
книги "Life at the Cell and Below-Cell Level..." |
|
{kind=link}