Life at the Cell and Below-Cell Level. The Hidden History of a Fundamental Revolution in Biology
"Dr. Ling is one of the most inventive biochemist I have ever met." |
Chapter 13. The Physico-Chemical Makeup of the Cell Membrane (p. 115-134) |
|
13.1 Background There is extensive evidence that a
(relative) diffusion barrier exists at the surface of most, if not all, living
cells [e.g., 4.1(1)]. In my view Pfeffer's Plasmahaut (protoplasm skin) is a more appropriate name for
this relative barrier. Yet the term, "cell membrane" has been so long
in usage that one can only accept it reluctantly—as one accepts reluctantly the
name, "cell." The key question we address in this chapter concerns
the physico-chemical makeup of this cell membrane. There are two opposing views
on the subject: one based on the membrane-pump-theory and one based on the LFCH (and AI Hypothesis). Following a brief description of both, I shall review
the results of experimental investigations aimed at finding out which model is
closer to the truth. (1)
The membrane theory Traube's study of the copper ferrocyanide
precipitation membrane launched the membrane theory. Unlike Thomas Graham's
starch-sized paper, but more like Abbe Nollet's pig bladder, copper-ferrocyanide membrane is not
only impermeable to colloids but also "impermeable" to crystalloids
like copper ion, ferrocyanide ion and sucrose. To explain this selective
permeability, Traube suggested in 1867 his atomic
sieve theory, in which pore size determines permeability or impermeability
—long before similar ideas were reintroduced again and again by a succession
of investigators including Michaelis,401 Mond
and Amson51 and Boyle & Conway.44
Traube's sieve idea proved wrong, however. Thus Bigelow, Bartell and Hunter demonstrated distinct osmotic activities
of copper-ferrocyanide membranes with pore diameters as wide as 0.5 micron
(5000 Ǻ).55; 56 р 96; 13 pp 656-657 A membrane with 5000 Ǻ-wide pores cannot act as a sieve to bar
the passage of (virtually impermeant)
"large" solutes like sucrose with a molecular diameter of only 8.8 Ǻ. However, earlier in 1855 L’Hermite
demonstrated how solubility (rather than pore size) may underlie
semipermeability or more correctly, differential permeability to various
solutes57; 56 p 98 (see also Liebig482). In support of
his solution theory, L’Hermite carried out an
ingenious and convincing experiment. He filled a cylinder with chloroform
(heaviest and stays at bottom), water and ether (lightest and stays at the top)
in that order. Now ether is (slightly) soluble in water; chloroform is not.
Accordingly, ether passes through the water layer to enter the chloroform
layer, but chloroform cannot go through the water layer to reach the ether layer.
Water in this case acts as a semipermeable medium, permeable to ether
but impermeable to chloroform. A well-known extension of L’Hermite’s
solution theory is Charles E. Overton's lipoidal membrane theory.21 Following an earlier
suggestion of Quinke, who postulated the existence of
an oil layer over the surface of living cells,542
Overton reiterated this postulation, adding that the different permeability of
the cell membranes to various non-electrolytes is attributed to their different
solubility in that membrane oil. Experimental support for Overton's lipoidal membrane
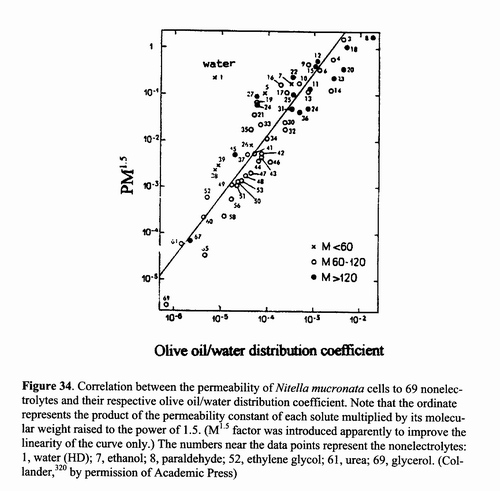
theory came from R. Collander320 and is shown as Figure 34. Here the
ordinate represents the product of experimentally measured permeability, P, of a particular substance
into Nitella cells multiplied by the molecular weight, M, of this
substance raised to the power of 1.5 (see legend for
justification). The abscissa represents the olive oil/water distribution
coefficient of the 69 compounds studied. The good correlation demonstrated suggests
that solubility in a lipoid cell membrane determines the relative permeability
of a nonelectrolyte. Behind this good appearance, there are serious unresolved
problems. Three will be mentioned.
Figure 34. Correlation between the permeability of Nitella mucronata cells to 69 nonelectrolytes and their respective olive oil/water distribution coefficient. Note that the ordinate represents the product of the permeability constant of each solute multiplied by its molecular weight raised to the power of 1.5. (M1.5 factor was introduced apparently to improve the linearity of the curve only.) The numbers near the data points represent the nonelectrolytes: 1, water (HD); 7, ethanol; 8, paraldehyde; 52, ethylene glycol; 61, urea; 69, glycerol. (Collander,320 by permission of Academic Press)
Firstly, the permeability was measured by
first exposing the Nitella cells to a solution containing the
nonelectrolyte under study for a length of time, before samples of the fluid of
the central vacuole (see Figure 1A) were collected and analyzed. Now, the
reader may recall that Höfler had demonstrated
in his later studies that the diffusion barrier to sucrose in the mature plant
cell he studied is the (inner) vesicular membrane (or tonoplast)
and not the (outer) plasma or cell membrane (Figure 1). Collander
did not address this problem. Therefore, a distinct possibility exists that the
diffusion barrier, which the data of Figure 34 represent, is not of the cell membrane
but that of the tonoplast. Tonoplasts
are special structures found in mature plant cells and cannot be seen as a
model of all cell membranes. Secondly, the distinguishing feature of a semipermeable
membrane is its overwhelmingly higher permeability to water than to solute
dissolved in water. Indeed, the first report on the subject by Abbé Nollet showed the pig
bladder to be permeable to water but impermeable to grain alcohol or ethanol.
Yet a look at Figure 34 reveals that the olive oil/water distribution coefficient
of ethanol is not lower than that of water but more than 100 times higher
than that of water. Thus the Nitella membrane should be 100 times more
permeable to ethanol than to water, contrary to facts underlying the very
concept of semipermeability. Thirdly, the interfacial tension of oil and water is
at least a hundred times higher than that actually measured at the cell surface
(see [13.4] for more details). Since both the atomic sieve model and the lipoidal
membrane model have encountered serious difficulties, a succession of
investigators attempted to combine the two models into one. Nathanson's mosaic membrane theory,438
Ruhland's ultrafilter
theory,422 Collander and Bärlund's lipoidal-filtration theory,403 Danielli-Harvey's paucimolecular
theory483 as well as the currently accepted Singer-Nicolson fluid-mosaic membrane200 (in which
phospholipid bilayer has replaced Overton's oil layer) are examples. In all
these models, a layer of oil or phospholipids forms a continuous barrier
between the cell interior and the external medium. In all these models the
cell membrane is a true membrane with a distinct boundary facing both inward
and outward. There is, however, one exception. That is the membrane model of
Wilhelm Pfeffer, often cited as the originator of the
membrane theory. Actually, in Pfeffer's model,
proteins make up a part or the entirety of the plasma membrane.18 p
156 And there may not be a distinct boundary separating the Plasmahaut from the cytoplasm.18 p 139 Therefore,
the (alleged) founder of the membrane theory has, in fact, much in common with
Franz Leydig's and Max Schultze's
thoughts on the chemical nature of the cell membrane [1]. To be continued |
|
|
Разделы книги
Contents (PDF
218 Kb) |
|
|
На страницу
книги "Life at the Cell and Below-Cell Level..." |
|
{kind=link}
{kind=link}