Life at the Cell and Below-Cell Level. The Hidden History of a Fundamental Revolution in Biology
"Dr. Ling is one of the most inventive biochemist I have ever met." |
Chapter 4. Evidence for a Cell Membrane Covering All Living Cells (p. 14-25) |
|
The
following is a summary of the evidence for the existence of a cell membrane
covering all living cells. 4.1 From studies of cell volume changes and solute permeability Cell volume change, the first subject
matter of cell physiological experimentations and observations, reflects
largely the movement of water in and out of cells. In the membrane theory,
water movement in turn reflects the permeability of the cell membrane to water,
and the permeability or impermeability to substances dissolved in water. (1)
A semipermeable diffusion barrier at the cell surface Chambers and Chambers showed that non-injurious dyes,
which could not penetrate the cell from the outside, diffused rapidly through
the cytoplasm when micro-injected into the cell interior. They stopped on
reaching the cell surface, unable to escape from the cell.22 (2)
Plasmolysis When a mature plant cell is exposed to a concentrated
solution of table salt (sodium chloride, NaCl) or cane sugar (sucrose), the
inside of the plant cell or protoplast (a name coined by von Hanstein,27
Figure 1B) shrinks away from the containing cell wall (Figure 1). The degrees of shrinkage varies with the strength of the NaCl or
sucrose solution used. This phenomenon was extensively studied by Hugo de Vries28
and is known as plasmolysis. To
explain the need for this tenet, consider the following. Many vertebrate cells
spend their entire life-spans—which may be as long as 100 years—in tissue
fluids containing NaCl as its main osmotically-active ingredient. If NaCl can
enter the cell, even slowly, the cell would sooner or later be chock-full of NaCl,
lose its osmotic equilibrium and probably die. Therefore, the cell membrane
must be impermeable to NaCl, permanently and absolutely. A
high point in the experimental study on plasmolysis
occurred in the year 1918, when K. Höfler
invented a method for measuring quantitatively the volume of the
(irregularly-shaped) shrunken protoplast29 (Figure 1 В). Applying
this method, he showed that in the mature cells of the plant, Tradescansia elongata, the
product of the volume of the protoplast, V, and the concentration of sucrose in
the bathing solution, C, is close to being a constant. This invariance of the
product, VC, was hailed as a decisive quantitative confirmation of the membrane
theory. It shows that the living cell indeed behaves like a perfect osmometer
in solutions containing different concentrations of an impermeant
solute like sucrose.30 Further
investigations, however, led Höfler to reverse
his earlier stand.31 Thus in later studies,
he discovered that it is not the whole cell enclosed by the cell membrane that
behaves like a perfect osmometer. Rather, it was only the central vacuole
enclosed by the vesicular membrane (or tonoplast),
which obeys the constant VC relationship: This new discovery poses a serious
difficulty for the membrane theory. At
high concentrations, sucrose causes the shrinkage of the central vacuole. This
is a fact. This shrinkage could not have happened without sucrose first
entering into the cell's cytoplasm, which completely surrounds the central
vacuole. Sucrose could not have entered the cytoplasm without first penetrating
the plasma or cell membrane, which in turn completely surrounds the cytoplasm.
Therefore, the cell membrane is not impermeable to sucrose—in contradiction to
the basic tenet that a solute that causes sustained plasmolysis
like sucrose is impermeant to the cell membrane. Other
studies showed that in concentrated solutions of an electrolyte (e.g., a salt
of potassium ion, K+), the surrounding cytoplasm may
actually swell even as the central vacuole is shrinking.32 This observation too
cannot be explained by the membrane theory. (3) Transient and sustained volume changes As I have just pointed out above, sodium chloride (NaCl)
is the major ubiquitous component of the tissue fluids of all vertebrates
including frogs and humans. Charles E. Overton—a distant cousin of Charles
Darwin—showed in 1902 that an isolated frog sartorius muscles—a thin and
flat muscle on the inside surface of the thigh, often highly developed in
tailors of early days (sartor, Latin)—retained their normal weights in a 0.7% NaCl
solution.25 Such a weight-preserving 0.7% NaCl solution was
described as isotonic by H. J. Hamburger; higher and lower
concentrations of NaCl solutions were respectively called hypertonic and
hypotonic26 When 5% methyl alcohol (methanol) was included in
an isotonic 0.7% NaCl solution, the muscle immersed in the solution showed no
change from its previous weight in the 0.7% NaCl solution alone. However, in a
solution containing 3% ethylene glycol and 0.35% NaCl solution, the muscle
shrank first followed by a slow return to a higher weight, close to that produced
by the unadulterated 0.35% NaCl solution alone. The explanations offered are as follows: Methyl
alcohol is, like water, highly permeable to the cell membrane. Hence, its
addition to an isotonic NaCl solution caused no change in the weight of the
muscle (if one assumes that the dilution of NaCl thus brought about is
trivial). Ethylene glycol is also permeant, but less
so than methanol. Therefore the inclusion of ethylene glycol caused first a
shrinkage followed by a return to normal and above normal weight as more and
more ethylene glycol enters the cells. In NaCl solutions at a concentration
higher than 0.7% or in a 0.7% NaCl solution containing also 3% glucose, the
muscle shrank and stayed shrunken. These observations were seen as once more
confirming the basic tenet of the membrane theory that only concentrated
solutions containing solutes like NaCl or glucose, which are absolutely and
permanently impermeant to the cell membrane can cause
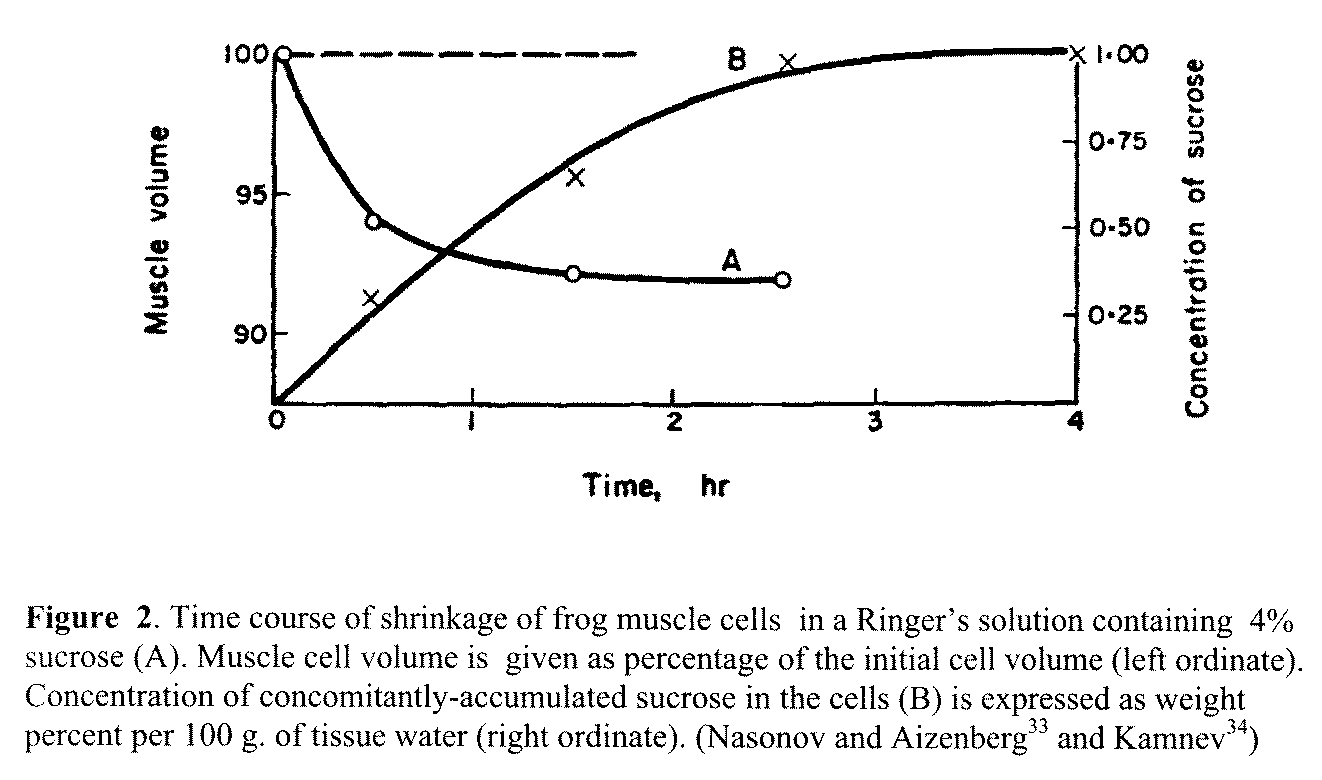
sustained cell shrinkage. Then something unexpected came up again. In 1937 and 1938, D. Nasonov,
E.I. Aizenberg33 and I. Ye. Kamnev34 from the former Soviet Union made
a simple discovery. They first showed that in a normal isotonic Ringer's
solution containing in addition 4% sucrose, frog muscle shrank until it reached
and then sustained a smaller volume (Figure 2)—as one would have expected on
the basis of the membrane theory. However, what one did not expect was that as
the muscle was shrinking, sucrose was actually entering into and
accumulating inside the shrinking cells. This inward permeation of sucrose
continued until a steady level—lower than that in the external medium—was
reached and then maintained (For additional details of their experiments, see
[8.2].)
With the advent of radioisotopes and associated
technology in the late 1930's and early 1940's, cell physiology entered a new
age of enlightenment.
Indeed with the radioactive tracer technique, (true) permeability—the sine que поп of the membrane
theory—can now be studied with precision and without ambiguity, often for
the first time {[13.3]; [15.2(1)]}. Thus the data shown in Table 1 once more confirm the existence of a diffusion barrier to labeled sucrose and NaCl as the surgical amputation of the frog muscle cells doubled the rate of penetration of sucrose into the muscle cells.23 However, the data also demonstrate that before the surgical amputation of the cell membrane, labeled sucrose could and did enter into (intact) muscle cells. Other studies have repeatedly and unequivocally established that solutes like sucrose23,35 and NaCl36 are permeant to the cell's membrane, confirming that permeant solutes can cause sustained cell shrinkage, a truth already suggested in Höfler's later discovery, and established by Nasonov, Aizenberg and Kamnev. And, in consequence, the elementary fact that living cell maintain steady and unchanging volumes in isotonic NaCl or sucrose solution can no longer be explained by the membrane theory. To be continued |
|
|
Разделы книги
Contents (PDF
218 Kb) |
|
|
На страницу
книги "Life at the Cell and Below-Cell Level..." |
|

{kind=link}
{kind=link}
{kind=link}